« Les Phytohormones » : différence entre les versions
| Ligne 407 : | Ligne 407 : | ||
--[[Utilisateur:Olivier.inf|Olivier.inf]] 6 mars 2009 à 22:43 (CET) | --[[Utilisateur:Olivier.inf|Olivier.inf]] 7 mars 2009 à 16:54 (CET)--[[Utilisateur:Olivier.inf|Olivier.inf]] 6 mars 2009 à 22:43 (CET) | ||
== L'éthylène == | == L'éthylène == | ||
Version du 7 mars 2009 à 17:54
Les hormones végétales
Le développement d’une plante ne se déroule pas au hasard mais à la fois de façon :
- Harmonieuse : une plante est généralement équilibrée dans ses proportions , la taille relative des différents organes est proportionnée le rapport surface aérienne surfaces parties souterraine demeure relativement constant.
- Coordonnée : apparition séquentielle d’organes. Une semence lors de sa germination émet d’abord une radicule qui pénètre dans le sol et fixe la jeune plantule qui va développer sa partie aérienne. (les fleurs apparaissent après les feuilles).
- Reproductible : pour une espèce donnée, si les conditions sont identiques les dimensions de l’individu arrivé à maturité sont comparables, les périodes de floraison ou de fructification se retrouvent à des époques comparables.
Le développement comprend une série d’événements au niveau cellulaire : division, élongation, différenciation, mort cellulaire qui sont intégrés à l’échelle du tissu et de l’organe via des interactions cellulaires générant en particulier des gradients morphogénétiques.
Le développement se déroule donc selon un plan propre à chaque espèce qui dans les conditions normales correspond à la mise en place séquentielle de programmes génétiques de développement se recouvrant partiellement. Par exemple, dans la floraison les gènes d’identité du méristème floral comme « leafy » interviennent dans la conversion méristème végétatif – méristème floral mais stimulent également l’activation des gènes d’identité d’organes floraux intervenant plus rapidement.
Dans le cas du développement végétal l’environnement a un poids particulier, une très forte influence sur le développement, Il s’agit des contrôle externes :
Les facteurs de l’environnement peuvent agir selon des effets que nous qualifierons de trophiques en conditionnant l’intensité du métabolisme cellulaire (T°, lumière,etc…), parfois selon des effets mécaniques (exemple vent). Et enfin selon une 3ème catégorie d’effets beaucoup plus subtile que nous appellerons effets signaux, une modification du milieu extérieur correspondant pour la plante à un signal qui va influencer son développement. Ces effets signaux peuvent faire intervenir les hormones comme intermédiaires ou agir après avoir été enregistré au niveau de la plante par des récepteurs capable de percevoir ces signaux et de les transformer en information utilisable par la plante. Les photorécepteurs comme le phytochrome représentent un exemple typique de perception de l’environnement lumineux et de contrôle de développement.
En ce qui concerne les contrôles internes :
Ils sont directement liés à la constitution génétique des individus, à leur génome qui contient une information de base (protéines enzymes facteur de transcription) et une information d’organisation susceptibles de percevoir, d’intégrer les signaux externes et de coordonner
Chez les végétaux, les hormones sont très souvent de petites molécules non protéiques. On connaît depuis longtemps 6 grandes classes d'hormones :
A l’heure actuelle chez les végétaux, on peut distinguer : Des hormones stimulatrices (qui induisent ou stimulent un phénomène physiologique) :
- Les Gibberellines
- Les Auxines
- Les Cytokinines
- Les Brassinostéroïdes
En parallèle on distingue des hormones à effets mixtes comme :
- l’acide abcissique
- l’éthylène
--Olivier.inf 6 mars 2009 à 17:13 (CET)
Les Gibbérélines
I- Historique :
Dès le début du siècle, des fermiers japonais avaient constaté que certains plants de riz étaient atteints de gigantisme. Ces plants cependant ne fructifiaient pas et ne présentaient donc pas d’intérêt pour la production.
- 1926 : KUROSAWA établit que cette anomalie dans la croissance résultait de l’infection par un Ascomycète parasite appelé Gibberella fujikuroi ainsi que le Fusarium moniliforme lorsque le champignon fut cultivé in vitro ( un extrait de son milieu de culture provoqua les mêmes symptômes d’élongation ).
- 1938 : on arriva à isoler de ces milieux de culture un mélange de substances actives appelées gibbérellines.
- 1956 : à partir d’une souche de Gibberella ne produisant qu’une seule gibbérelline on put isoler et caractériser chimiquement l’acide gibbérellique ou GA3 (travaux de CROSS).
Pendant ce temps, les physiologistes démontraient les effets spectaculaires des gibbérellines isolées des filtrats de cultures de champignons sur la croissance de végétaux (à très faibles doses ces substances stimulent en particulier la croissance des espèces naines : haricot, pois, maïs).
I- Définition des gibbérellines :
Les gibbérellines sont une famille de phytohormones. Le composé actif est appelé acide gibbérellique. Les gibbérellines sont nommées G ou Ga suivi d'un chiffre (de 1 à 125). La Ga3 est la mieux connue.
Ce sont des substances synthétisées par les plantes possédant le squelette gibbane et actives vis-à-vis de tests biologiques spécifiques tels que la croissance de mutants nains (Maïs) ou la production d’αamylase par des albumens d’orge.
125 variétées, 10 chez les champignons, 100 chez les végétaux et 15 communes. Seules les GA1 et GA3, ont une forme suffisament active, pour la culture du cannabis.

II- Biosynthèse et Métabolisme des Gibberellines :
Les gibbérellines appartiennent au groupe des terpénoïdes composés résultant de la condensation d’unités isoprène elles-mêmes provenant d’unités acétate.
Acétate ==> isoprène ==> terpènes
Les terpènes constituent entre autre le principe odoriférant des végétaux. Cette odeur est due à la libération des molécules très volatiles contenant 10, 15, 20 atomes de carbones. Extraites ces molécules sont employées comme condiment (girofle) ou comme parfum (rose, lavande). Nombre d'entre eux possèdent de propriétés antiseptiques, d'où divers emplois dont l'embaument qui est resté dans le terme balsamique donné aux plantes et aux huiles qui en sont tirées.
Formule des terpènes :

III- Répartition et Transport des Gibberellines dans la plante :
Les gibbérellines sont présentes chez toutes les plantes supérieures, elles sont synthétisées également par certains champignons. Les gibbérellines détectées varient selon le stade de développement.
On pense que les sites de synthèse sont les organes contenant les concentrations les plus élevées en gibbérellines, apex des tiges et des racines, jeunes feuilles, mais aussi embryon et tissu de réserve des graines en développement, fruits…
Les concentrations habituelles sont de 0,1 à 100 ng / g de tissu frais mais de 1 à 10 μg au niveau des graines.
Les gibbérellines ne présentent pas de transport polarisé à la différence de l’auxine. Appliquées à un niveau quelconque de la plante, elles peuvent avoir des effets régulateurs sur toutes les autres parties. Elles ont été retrouvées dans la sève brute et la sève élaborée et leur vitesse de transport (5 cm/h) analogue à celle des sucres laisse supposer qu’elles sont transportées passivement dans les flux de sève dans le xylème et le phloème.
Un transport de cellules à cellules de type symplastique ( Déf : Le symplasme désigne le continuum intracellulaire formé par les cellules végétales par le biais des plasmodesmes. Les cytoplasmes de cellules ainsi reliés ne forment alors qu'un seul compartiment partagé par toutes les cellules. La taille des plasmodesmes régule la taille des solutés pouvant naviguer activement depuis une cellule vers une autre. Dans les racines, le symplasme assure le passage de l'eau et des solutés du sol vers le cylindre central où siègent les faisceaux conducteurs de sève )est également probable.
IV- Effets Physiologiques :
Au niveau cellulaire comme les auxines, les gibbérellines ont à la fois une action sur la division, l’élongation et la différenciation. Parmi les effets observables on peut citer :
- L’action sur la croissance des tiges (au niveau des racines et feuilles on observe de très faibles réponses);
- Croissance des fruits effet commun avec les auxines, mais les gibbérellines agissent sur des espèces pour lesquelles l’auxine n’a pas d’action (Rosacées, Pêcher, Pommier, Raisins). La parthénocarpie ( du grec "graine vierge" est la production de fruits sans fécondation d'ovule ) peut être obtenue avec des gibbérellines;
- Levée de dormance. L’application de gibbérellines à des bourgeons dormants permet la levée de dormance et leur débourrement. Même effet sur la levée de dormance des graines.
- Initiation de la floraison. Pour des espèces ayant des exigences photopériodiques ou de vernalisation pour fleurir, la transformation d’un méristème végétatif en méristème floral peut être obtenue dans de nombreux cas par application de gibbérellines. Sans que l’on sache si ces hormones sont directement impliquées dans le processus physiologique normal.
L'apport exogène de Calcium est IMPERATIF pulvérisé ou dans la solution nutritive
--Olivier.inf 6 mars 2009 à 18:41 (CET)
Les auxines
I- Historique :
- 1880 : Observation (Darwin) du phototropisme chez les coléoptiles de graminées
excitation perçue au sommet et transmise vers la base
- 1926 : Récupération par diffusion d’une substance active sur la croissance
appelée auxine, d’auxein = croître. Mise au point d’un test biologique permettant d’apprécier les teneurs en substance active.
- 1934 : Identification chimique de l’auxine à l’acide ß indolacétique (isolé
initialement à partir d’urine humaine) puis caractérisation de cette structure dans les tissus végétaux (Zea mays) par Haagen – Smith en 1942.
- 1925-1970 : Recensement des différentes réponses des plantes à l’action de l’AIA. Caractérisation de substances naturelles ou synthétiques à action auxinique.
- 1970-2009 : Etudes sur le mode d’action de l’AIA au niveau moléculaire en particulier dans le phénomène de grandissement cellulaire.
II- Définition :
C'est une hormone végétale stimulatrice (qui induit ou stimule un phénomène physiologique). Au sens strict, l'auxine est de l'acide indole-acétique (AIA). Le terme d'auxines a ensuite été élargi à un ensemble de substances possédant des propriétés physiologiques voisines et une conformation chimique apparentée ( Acide Indole Butyrique, Acide Naphtalène Acétique et l'Acide Phényl Acétique). L’AIA et les autres auxines semblent présentes chez toutes les plantes vasculaires.
Représentation de la molécule d'AIA et sa nature chimique :

III-Lieu de synthèse et transport de l'auxine :
La synthèse de l'auxine s'effectue dans les apex des tiges, dans les méristèmes et jeunes feuilles des bourgeons terminaux.Ceux-ci reçoivent les précurseurs, comme le tryptophane (Le tryptophane est un acide aminé, qui dans sa configuration lévogyre (L-tryptophane) est l'un des 20 acides aminés constituant des protéines), qui eux sont fabriqués dans les feuilles plus âgées, à la lumière. Les méristèmes intercalaires sont également des lieux de synthèse très actifs.
La migration (à distance du lieu de synthèse) est primordiale dans le cas de l'auxine : synthétisée dans les apex et les entre-noeuds des tiges et des rameaux, cette hormone doit être distribuée dans tous les tissus, y compris les racines, où elle s'accumule. L’auxine comme les autres hormones doit pour jouer son rôle de messager chimique ne pas demeurer à une concentration constante dans les tissus mais voir ses teneurs fluctuer. C’est la « vague auxinique » qui va déterminer une réponse dans le temps, adaptée au contexte de morphogenèse et de la différenciation. L'auxine se déplace préférentiellement dans le phloème (Le phloème est le tissu conducteur de la sève élaborée qui est une solution riche en glucides tels que le saccharose, le sorbitol et le mannitol chez les plantes vasculaires. Le phloème a aussi un rôle de réserve avec les parenchymes et un rôle de soutien avec les fibres libériennes. Dans les troncs des arbres, le phloème peut faire partie de l'écorce, pour cette raison son nom provient d'un dérivé du mot grec phloos qui signifie écorce).

Sa conduction est polarisée : elle s'effectue plus facilement de l'apex vers la base de l'organe.
Ce transport polarisé de l'auxine :
- a des conséquences sur l'organogenèse : il y a différenciation de racines à la base et de bourgeons à l'apex, en relation avec l'action rhizogène de l'auxine à forte dose et avec l'action de l'auxine à faible dose sur la différenciation des bourgeons
- permet de comprendre les tropismes : par exemple, dans le contexte du phototropisme, la lumière provoque (par un mécanisme que l'on ignore) une migration d'auxine depuis la face éclairée vers la face sombre, engendrant ainsi un allongement de la face sombre lié à l'effet de l'auxine à forte dose sur l'élongation cellulaire.
IV-Rôles de l'auxine dans la morphogenèse végétative :
L'auxine et l'élongation cellulaire :
L'auxine est la principale hormone agissant sur l'augmentation de la taille des cellules. Cet effet, qui dépend des concentrations intracellulaires d'auxine et de la nature des organes, s'exerce sur des cellules jeunes en cours d'élongation, au moment où la paroi est extensible.
On sait que l'élongation cellulaire est un processus complexe qui fait intervenir une absorption d'eau, l'extension de la paroi sous l'effet de la turgescence, et l'incorporation de nouveaux composés entre les mailles de fibrilles de cellulose ainsi distendues.
L'auxine agit en fait sur l'élongation cellulaire à deux niveaux :
- d'une part au niveau de la paroi, dont elle provoque le relâchement.Dans le détail, l'auxine stimule au niveau de la membrane plasmique une pompe à protons, entraînant ainsi une acidification du milieu : le pH au voisinage de la paroi tombe de 6.5 à 4.5. L'efflux de protons a plusieurs conséquences, toutes favorables au relâchement de la paroi : rupture de liaisons acidolabiles entre l'extensine, les hémicelluloses et composés pectiques, et la cellulose ; déplacement du calcium qui soudait entre elles les chaines uroniques des composés pectiques ; entrée d'ions K+ provoquant conjointement une entrée d'eau d'où une augmentation de la turgescence cellulaire ; activation de certaines enzymes, de type cellulases et protéases, susceptibles d'hydrolyser les composés de la paroi.
- d'autre part sur les synthèses protéiques[/i], en modifiant l'expression génique. Il est établi que l'auxine agit sur l'activité génique en régulant la synthèse d'ARN codant pour des protéines nécessaires à l'élongation. Ces protéines spécifiques de l'élongation cellulaire n'ont pas été clairement identifiées à ce jour. Il reste également à préciser leur rôle et le lien qu'elles peuvent avoir avec la stimulation de l'efflux de protons.
L'auxine et la division cellulaire:
L'auxine stimule les mitoses (division cellulaire), mais cette action ne s'exerce pas indistinctement sur tous les méristèmes : l'auxine n'agit pas (ou peu) sur la prolifération au niveau des méristèmes primaires. En revanche, elle a une action très marquée sur la prolifération des cambiums (en vert sur le schéma ci-dessous) :

Aspects macroscopiques des rôles de l'auxine:
- L'auxine et la croissance des organes végétatifs :
L'auxine contribue à la croissance des tiges et des rameaux, à partir des bourgeons apicaux ou axillaires. Son action sur la croissance en longueur dans la zone d'élongation subapicale est maximale pour de concentrations en auxine relativement élevées. Par contre, l'élongation des entre-noeuds n'est pas le fait de l'auxine. Au niveau des feuilles, les pétioles et les gaines ont leur élongation stimulée par l'auxine. Les limbes des feuilles de Monocotylédones ont leur croissance stimulée par l'auxine, tandis que celle des limbes de Dicotylédones est inhibée par l'auxine. L'action de l'auxine sur l'élongation des racines est toute différente de son action sur les tiges. Elle se ramène à un effet inhibiteur aux concentrations moyennes.
- L'auxine et l'organogenèse :
La néoformation (ou différenciation) des bourgeons est induite par les cytokinines, sous réserve de la présence de faibles doses d'auxine. Pour des concentrations plus fortes, l'auxine inhibe la différenciation des bourgeons. À forte dose, l'auxine inhibe également le débourrement des bourgeons. L'un des effets organogènes le plus marquant de l'auxine est son pouvoir rhizogène : appliquée à de concentrations assez fortes, l'auxine provoque l'apparition de racines.
--Olivier.inf 6 mars 2009 à 16:52 (CET)
Les cytokinines
I- Historique :
- 1941 : Van Overbeek met en évidence les propriétés actives du lait de noix de coco, vis-à-vis de la croissance de jeunes embryons de Datura stramonium. Ce milieu est toujours utilisé en culture de tissus végétaux.
- 1954 : Le groupe de SKOOG' montre que la croissance in vitro des tissus de moelle de tabac ne peut se faire avec la seule présence d’auxine (faible croissance pas de division, seulement grandissement cellulaire). La recherche de substances actives conduit à mettre en évidence l’action positive du lait de noix de coco, de l’extrait de levure et d'ADN autoclavé.
- 1955 : Miller obtient à partir de sperme de Hareng autoclavé (très riche en ac. nucléïques) une substance capable d’induire la division cellulaire des tissus de moelle de tabac à de très faibles concentrations 1 μg/litre.
Cette substance a été identifiée il s’agit de la 6-furfurylaminopurine ou kinétine actuellement encore utilisé comme régulateur de croissance.
D’autres substances synthétiques de nature voisine et des dérivés de l’adénine isolés des végétaux ont une action comparable. L’ensemble de ces substances est regroupé sous le terme de cytokinines.
II- Définition :
Les cytokinines sont des substances proches des bases puriques, (adénines substituées).
III- Biosynthèse et Métabolisme des cytokinines :
A côté de la kinétine d’autres substances synthétiques à activités cytokinine existent dont la plus connue est la 6-benzyladénine, la Zéatine.
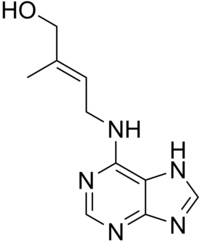
Les cytokinines sont des constituants naturels des végétaux qui interviennent dans la synthèse des acides nucléiques. Les cytokinines naturelles connues résultent de la substitution d’un hydrogène du NH2 (en 6) par une chaîne, à 5 atomes de carbone correspondant à une unité isoprène de type pyrophosphate d’isopentényle. Les cytokinines existent et sont actives à la fois à l’état libre à l’état de nucléosides et de nucléotides.
IV- Répartition et Transport des Cytokinines dans la plante :
Il est classiquement admis que les cytokinines sont produites de façon préférentielle dans les racines, bien que les embryons, les jeunes fruits, les bourgeons aient aussi une autonomie de production. Elles sont présentes dans les racines en grande quantité et sont synthétisées à partir de précurseurs radioactifs. On retrouve des cytokinines dans les exsudats racinaires de certaines plantes (Maïs). Les feuilles sont dépendantes des racines pour la production de cytokinine. Les cytokinines seraient transportées dans le xylème. Appliquées de façon exogène au niveau des feuilles elles migrent peu. Un des effets des cytokinines est de permettre 'la cytokinèse c'est-à-dire la formation d’une paroi transversale assurant la séparation de deux cellules filles. C’est en raison de cette action spécifique sur cette phase de la division cellulaire que le nom de cytokinine a été donné à ces hormones. Il faut également remarquer que dans les conditions des essais biologiques les cytokinines seules sont sans action sur la division cellulaire mais qu’elles ne peuvent agir qu’en présence d’auxine. Ceci est un exemple de complémentarité d’action entre deux substances de croissance => synergie qui doit vraisemblablement se retrouver dans de nombreux cas. Les cytokinines permettent la différenciation de bourgeons sur des tissus en culture leur action est contrebalancée par celle des auxines qui favorisent la production de racines, la différenciation du tissu dépendant en fait de l’équilibre auxines/cytokinines. La formation de tiges est favorisée par un apport élevé en cytokinines et un apport faible en auxines. La formation de racines est favorisée par un apport faible en cytokinines et un apport fort en auxines.
V- Effets Physiologiques sur la plante cannabis :
- Retardent la sénescence foliaire;
- Associé à l'auxine, activent la division cellulaire (Accélèrent la séparation des gênes dans le noyau nuclèïque);
- Accélèrent le déploiement des pales des feuilles;
- Augmentation de l'absorption de l'eau et des nutriments.
- Accélèrent la mise à disposition des oses (par le Xylème) au niveau de la plante;
- Production accrue de la chlorophylle.
--Olivier.inf 6 mars 2009 à 21:30 (CET)
Les brassinostéroïdes
I- Historique:
- 1968 : Ces molécules ont été initialement isolées avec du pollen de Brassica majus sous le terme de brassines.
- 1979 : Mise en évidence d'une molécule spécifique de ce groupe appelé Brassinostéroïde.
II- Définition :
Les Brassinosteroides (BRs) ou brassinolides sont un groupe d’hormones steroïdiennes végétales. Leurs actions se produisent au niveau d'un photorécepteur à la lumiére bleue soit les cryptochromes. Une quarantaine de structure actives ont été actuellement caractérisées les Brs étant présents chez les algues, fougères, gymnospermes, angiospermes mais pas chez les microorganismes. Le brassinolide est le plus actif biologiquement et le plus répandu. Ces molécules appliquées sur divers systèmes expérimentaux à des concentrations nanomolaires présentent un effet marqué sur l’élongation cellulaire ou sur la prolifération cellulaire.
Formule des Brassinolides:

III- Biosynthèse des Brassinostéroïdes
Le brassinolide (Br type) est un stéroïde présentant un squelette cholestane qui possède un cycle B-7oxalolactonique et 2- hydroxyles adjacents sur le cycle A(C₂α et C₃α) et sur la chaîne latérale C22 et C23.
La voie de synthèse du Brassinolide à partir du campestérol, un stérol végétal de répartition très générale a été établie au cours des dernières années.
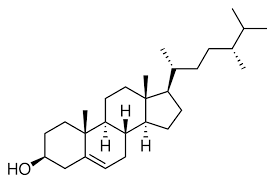
IV- Effets physiologiques des brassinostéroïdes :
Les brassinostéroïdes ont des effets pléiotropiques sur les systèmes végétaux. Leur action est située au niveau de la mitose et l’élongation cellulaire. Ils interviennent également dans la différenciation des tissus vasculaires. Ils accélèrent la sénescence dans des systèmes simplifiés (feuilles, cotylédons isolés) par des effets antagonistes des cytokinines. Les Brs contrôlent l’expression génique en favorisant la transcription de certains gènes, c'est à dire qu'ils permettent la copie de l'ADN en ARN, ou de l'ARN en ARN messager. C'est la première étape du processus qui permet de passer de l'ADN à la protéine, ou plus concrêtement du gène à son produit. Ils ont aussi une action d'accélération du mouvement de la séve élaborée ou brute donc par effets redondants une meilleure absorption des nutriments et permettent aussi une amélioration des phénomènes osmotiques. Ils luttent contre le froid, la chaleur, ils ont aussi une action détoxifiante (sels toxiques), aide à une germination plus rapide des graines et les brassinolides ont un effet potentialisateur de la rhizogénèse. Enfin, ils aident la plante à se défendre de toute attaque par des agents pathogènes tels que les bactéries, champignons etc... L'apport exogène de calcium est IMPERATIF sous peine de sévères carences calciques.
L'Acide abscissique
I-Historique:
- 1960 : WAREING et ses collaborateurs au Pays de Galles recherchaient la cause de l’arrêt de la croissance des arbres en automne et le facteur qui provoque la formation des bourgeons dormants. Ils obtinrent à partir des feuilles d’Acer pseudoplatanus un extrait acide qui était un puissant inhibiteur de la croissance et qui appliqué aux apex des tiges feuillées était capable d’induire la formation de bourgeons dormants. Ils appelèrent la substance active encore inconnue : la dormine.
- 1966 : L’isolement de la dormine par CORNFORTH et al. permet sa caractérisation chimique et fut suivie par des travaux qui montrèrent que la dormine est la substance qui inhibe de croissance du Lupin.
- 1967 : Découverte de l'abcissine II c'étaient en fait la même substance qui fut définitivement appelée acide abscissique(ABA).
II- Définition de l'acide abscissique :
L'acide abscissique ou ABA (de l'angl. abscissic acid) est un sesquiterpénoïde composé de 15 carbones de formule C15H20
O4.
III- Nature chimique et Biosynthèse :
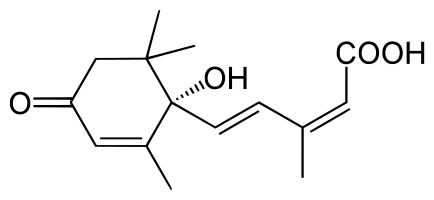
Deux voies de biosynthèse ont été successivement proposées pour l’ABA, la première dite voie en C15₅ correspondrait à la condensation de 3 molécules d’isopentenyl pyrophosphate selon un mécanisme analogue à celui de la synthèse des gibberellines.
La 2ème voie dite en C40₀ a été caractérisée plus récemment elle correspond à une coupure de caroténoïdes en C40₀ du type zeaxanthine selon la séquence :
Zéaxanthine ==> Violaxanthine==> Xanthoxine==> ABA aldéhyde ==> ABA
Formule de la Zéaxanthine :
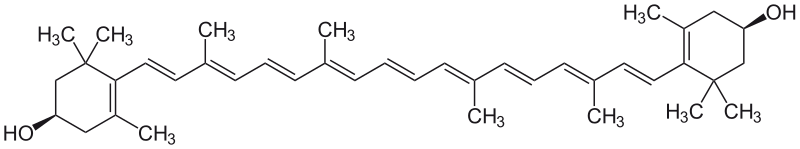
La zéaxantine C40H56O2 est un pigment de la famille des xanthophylles (caroténoïde) qui donne sa couleur jaune aux grains de maïs.
Effets physiologiques et mécanismes d’action de l'ABA :
- Action sur la fermeture des stomates:
Il s’agit d’un phénomène très important au plan physiologique puisqu’il permet de contrôler les pertes d’eau de la plante et de maintenir l’homéohydrie. C’est un exemple de réponse rapide à une hormone de l’ordre de quelques minutes (lors de l’apport d’ABA exogène). Harris et coll (1990) en utilisant un radio immuno essai (extrêmement sensible) pour l’ABA ont été capables de quantifier l’ABA dans une seule cellule de garde. A la suite d’un stress hydrique on observe un accroissement du taux d’ABA par un facteur 20. La production d’ABA se ferait en 1er au niveau des racines stressées qui perçoivent le stress et l’ABA serait transporté vers les apex.
- Formation des graines et dormance :
L’ABA intervient comme nous le verrons ultérieurement dans le contrôle de l’expression de gènes qui correspondent à des protéines de réserve des graines et à des protéines permettant sans dommage la déshydratation des tissus (les déhydrines). Parmi ces protéines une classe particulière a été spécialement étudiée ce sont les LEA protéines (late embryogenesis abundant) produite durant les phases tardives de l’embryogénèse. Leur expression est associée à l’acquisition de la tolérance à la déshydratation et elles sont censées protéger les structures cellulaires des effets de la perte d’eau (protection de protéines ou de membranes). L’ABA est, par ailleurs, nécessaire à l’entrée en dormance des graines et des bourgeons. L’ABA est d’une manière générale un antagoniste des gibbérellines dans des phénomènes comme la dormance ou la production d’α-amylase par les cellules d’aleurone.
- L'Abcission :
Bien que l’hormone ait été initialement caractérisée en relation avec l’abcission. Ce sont des doses supraphysiologiques qui sont actives et on pense que ces doses entraîneraient la surproduction d’éthylène véritable hormone responsable de l’abcission.
En conclusion le nom de dormine aurait été beaucoup plus adapté pour ce que nous appelons aujourd’hui l’acide abcissique.
--Olivier.inf 7 mars 2009 à 16:54 (CET)--Olivier.inf 6 mars 2009 à 22:43 (CET)
L'éthylène
I- Historique :
- 1886 : NELJUBOW une jeune Botaniste russe observait l’effet du gaz d’éclairage sur la morphologie de plantules de pois : raccourcissement et épaississement des tiges, perte du géotropisme négatif : ensemble de réponses regroupées sous le terme de triple réponse. Parmi les différents effets de l’éthylène ce sont cependant les observations relatives à la maturation des fruits qui ont été décisives dans la découverte de son rôle hormonal.
- 1924 : L’éthylène permet le jaunissement et la maturation des citrons.
- 1937 : On découvre que les émanations gazeuses de pommes mûres initient la maturation des fruits verts et que l’éthylène constituait le gaz actif (première démonstration de la production d’éthylène par un végétal). A partir de ce moment on attribue un rôle à l’éthylène dans la maturation des fruits et l’on montre que de nombreux fruits émettent de l’éthylène.
- 1955-1960 : le développement de la chromatographie en phase gazeuse fit franchir une nouvelle étape car cette méthode très sensible et particulièrement adaptée à la détection de ce gaz permet de montrer que l’éthylène était présente dans toute les parties de la plante.
Parallèlement on démontrait au-delà de la maturation les actions diverses de l’éthylène sur le développement des végétaux.
- 1969-2009 : Ce composé était finalement rangé parmi les hormones végétales. Produite par les végétaux, active à faible dose et à distance du lieu de synthèse l’éthylène répond tout à fait à la définition d’une hormone.
Elle représente cependant des caractéristiques particulières au niveau du transport : [b][i][u]on observe en effet une diffusion gazeuse à l’intérieur de la plante mais aussi à l’extérieur d’où la possibilité d’action sur d’autres individus.
II- Définition :
C'est un gaz et aussi une phythormone en botanique. C'est la seule phythormone pour laquelle on connaît le récepteur. L'éthène (éthylène) est le plus simple des alcènes. L'éthylène à pour formule à l'état gazeux H2C=CH2. La production permanente d'éthylène semble être necessaire au développement normal des plantes supérieures.
Formule de l'éthylène :

Depuis longtemps il avait été démontré que la méthionine (ac. aminé) était un précurseur de l’éthylène. En effet, si on apporte de la méthionine marquée à des tranches de pommes ou de bananes on observe une incorporation de la radioactivité dans l’éthylène. L’éthylène dériverait des carbones 3 et 4.
IV- Effets physiologiques :
L’éthylène peut être considéré comme une hormone mixte avec des effets positifs tels que l'initiation de la floraison, abscission, sénescence ainsi que la germination et des effets négatifs sur le développement en inhibant la croissance des végétaux.
Elle exerce une influence sur toutes les phases du développement de la germination à la sénescence souvent en interaction avec d’autres hormones.
Sans entrer dans le détail de ces modifications il faut signaler qu’elles sont précédées chez de nombreux fruits par un accroissement très net de l’intensité respiratoires que l’on appelle crise climactérique( Déf : Un fruit est dit "climatérique" ou "climactérique" d'après le terme anglais si sa maturation est dépendante de l'éthylène et associée avec une augmentation de la respiration cellulaire des tissus. Les fruits climactériques les plus connus sont : la banane, la pomme, la poire, la tomate, le melon, la pêche, l'avocat...Par opposition, un fruit non-climactérique est un fruit dont la maturation est indépendante de l'éthylène et non associée à une augmentation de la respiration des tissus. Les fruits non-climactériques les plus connus sont : les agrumes, le raisin, la fraise, la cerise...) la période antérieure ou phase pré climactérique étant une période d’activité métabolique ralentie.
La production d’éthylène est très sensible aux facteurs de l’environnement : lumière, température, différents types de stress (blessures, radiations, sécheresse, attaques par les microorganismes, etc…). Dans le cas de ces agressions cette synthèse accrue d’éthylène s’accompagne de la formation de composés phénoliques, les enzymes de synthèse PAL ou d’oxydation (peroxydase) de ces composés étant nettement activées. L’éthylène déclenche ainsi des réactions de la plante qui peuvent être assimilées à des sortes de réactions de défense(cicatrisation, protection…) d’où l’appellation d’Hormone de Stress.
La production d’éthylène est stimulée par les auxines (naturelles ou synthétiques). Les travaux d’ ABELES et de BURG (1968-1972) ont montré que de nombreuses réponses obtenues chez les plantes lors de l’application d’auxine pouvaient être reproduites par l’exposition des plantes à l’éthylène. Ainsi de nombreuses réponses attribuées à l’auxine aux fortes concentrations se produiraient par l’intermédiaire de l’éthylène (inhibition de l’élongation). Cette interaction pourrait fournir un contrôle naturel lors de la production excessive d’auxine.
V- Voies de biosynthèse et régulation de la synthèse :
Changements biochimiques lors de la maturation :
- Hydrolyse des composés pectiques => pectine soluble,
- Hydrolyse de l’amidon => sucres,
- Disparition des acides organiques => oses,
- Disparition des substances astringentes tels que les tannins.
Les changements biochimiques lors de la maturation s'opérent selon différentes étapes intermédiaires ont été caractérisées selon la séquence ci-dessous :
Méthionine => S-adenosyl méthionine (SAM)=> Acide cyclopropane carboxylique (ACC)=> Ethylène

La méthionine joue un rôle particulier dans la biosynthèse des protéines, puisque toutes les chaînes protéiques démarrent par l'incorporation d'une méthionine en position N-terminale. D'autres résidus méthionine peuvent ensuite être incorporés de manière interne à la chaîne polypetidique. La première méthionine des protéines n'est pas toujours retrouvée dans les protéines terminées. Elle est en effet fréquemment clivée par une enzyme spécifique appelée méthionine aminopeptidase.
La vitamine B12 ainsi que la vitamine B9 agissent en tant que co-enzyme dans la synthèse de méthionine. En effet, c'est sous la forme de l'enzyme méthyl B12 qu'un groupement méthyl est ajouté à l'homocystéine pour former la méthionine.
On a pu montrer que l’apport de l’éthylène déclenche la crise climactérique et les phénomènes de maturation qui s’en suivent.
Des mesures de la production d’éthylène dans le fruit révèlent que la quantité de gaz s’accroît avec la crise climactérique.
Selon les fruits on constate que la production d’éthylène est parallèle à la montée de la crise climactérique ou la précède en revenant à sa valeur initiale lors de la montée (Banane).
On considère donc que l’éthylène est l’hormone de maturation naturelle des fruits.
La maturation des fruits peut être considérée comme une étape précoce de la sénescence qui est définie par rapport à des critères de consommation. L’éthylène de façon plus générale, induit la sénescence chez d’autres organes comme les fleurs ou les feuilles.

--Olivier.inf 6 mars 2009 à 16:27 (CET)